Lorsque la première barrière de l'immunité innée s'avère insuffisante, une immunité dite adaptative intervient chez les vertébrés.
Les cellules immunitaires ont pour origine la moelle rouge des os.
Parmi les cellules intervenant dans l'immunité, nous retiendrons les lymphocytes B et T.
Les lymphocytes B (de bone, os en anglais) sont capable d'assurer leur rôle des la sortie de la moelle.
Les lymphocytes T (de thymus) ne peuvent assurer leur rôle qu'après être passé par le thymus, une glande située à la base du cou. Le thymus est un des organes lymphoïdes dans lesquelles se situent les lymphocytes.
Les lymphocytes peuvent être observés dans un frottis sanguin, mais leur détermination est difficile.
Intervention des lymphocytes B ou immunité à médiation humorale
La première étape de la réponse immunitaire adaptative est une reconnaissance de l'antigène par les lymphocytes B.
La reconnaissance de l'antigène par l'anticorps est du modèle clé/serrure. c'est une complémentarité de forme de l'anticorps par rapport à l'antigène qui permet la reconnaissance. C'est le lymphocyte porteur de l'anticorps complémentaire qui est sélectionné.
Une électrophorèse des protéines sériques (du sérum), permet de déterminer de quelle nature chimique sont les anticorps fixés sur les LB.
On constate que ce sont les gammaglobulines qui différencient le sérum d'un malade de celui d'un sujet sain. On appelle encore ces anticorps des immunoglobulines.
La structure des immunoglobulines est la suivante :
Une immunoglobuline est constituée de deux chaînes protéiques lourdes et de deux chaînes protéiques légères liées par des ponts disulfures. Les régions variables peuvent fixer deux antigènes par anticorps selon le modèle suivant :
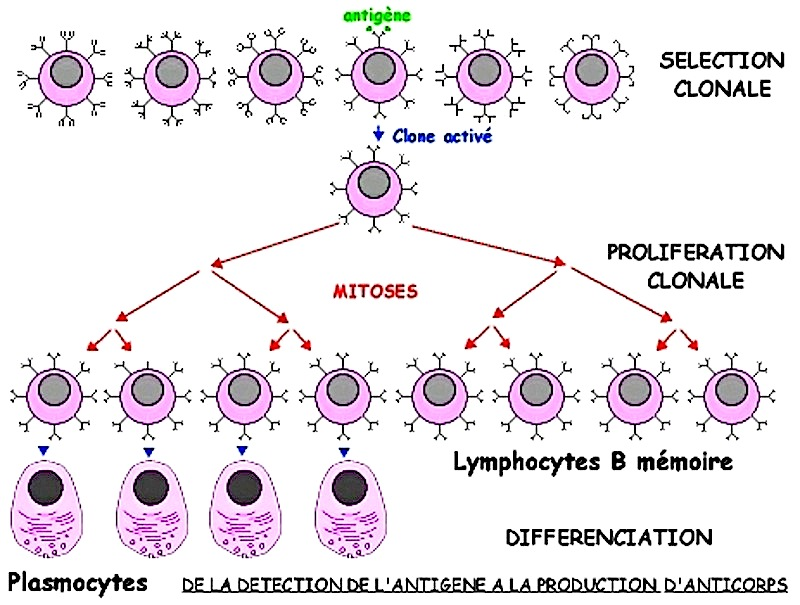
Lorsque le lymphocyte B porteur de l'anticorps anti-antigène a été sélectionné, il se met à se multiplier par mitoses. Après la phase de sélection clonale, il y la phase de prolifération ou amplification clonale.
Ce n'est qu'après cette phase que les LB commencent à se différencier en LB mémoire et enplasmocytes.
Les deux électronographies suivantes permettent de comparer les lymphocytes B et les plasmocytes.
Les plasmocytes sont des cellules de beaucoup plus grande taille que le lymphocyte B. La présence de mitochondries indique que c'est une cellule très active. La présence d'une grande quantité de réticulum avec ribosomes permet de constater que la cellule fabrique de grandes quantités de protéines, les anticorps. Chez le plasmocyte, les anticorps ne restent pas attachés au plasmocyte mais sont libérés dans le sang. On parle d'anticorps circulants.
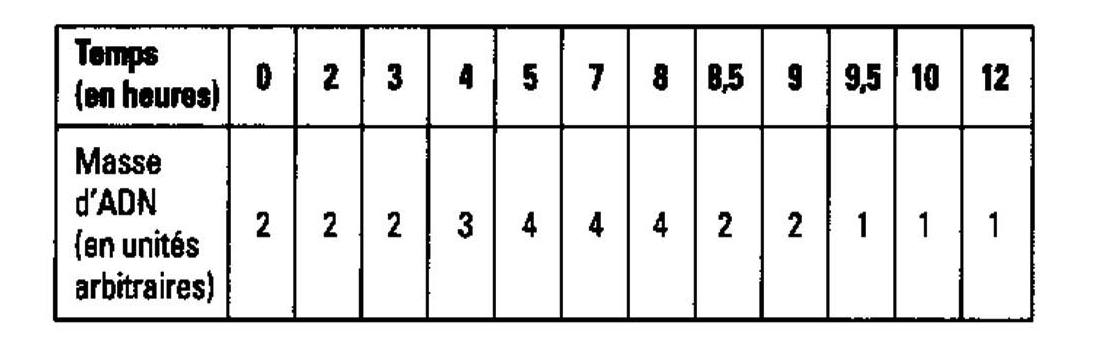
L'expérience dite de la chambre de Marbrook permet de comprendre comment la différenciation des LB en plasmocytes a lieu.
Les résultats de cette expérience permettent de comprendre que la production d'anticorps s'effectue bien mieux en présence de lymphocytes T4 (actuellement appelés LT CD4). Il y a donc une substance sécrétée par les LT CD4 qui accélère la différenciation des lymphocytes B.
Il s'écoule un certain temps entre le moment où les antigènes se développent et celui où les anticorps sont libérés dans le sang. A partir de ce moment-là, il y a séropositivité.
Les anticorps libérés par les plasmocytes constituent avec les antigènes, des structures appeléescomplexes immuns qui permettent la fixation des antigènes.
Schéma d'interprétation d'un complexe immun
L'élimination des complexes immuns se fait par l'intervention des macrophages qui les phagocytent tout en conservant les déterminants antigéniques sur leurs surfaces membranaires. Il y a donc une véritable coopération cellulaire entre les plasmocytes et la macrophages.
Intervention des lymphocytes T ou immunité à médiation cellulaire
Les lymphocytes B peuvent intervenir sur des antigènes libres dans l'organisme comme des bactéries. En revanche, si les antigènes s'installent dans des cellules, ou si des cellules sont transformées (par exemple cancérisées), les lymphocytes B ne peuvent pas intervenir. C'est le rôle des lymphocytes T.
Nous retiendrons deux types de lymphocytes : les lymphocytes T CD8 (anciennement lymphocytes T8) et les lymphocytes T CD4 ( lymphocytes T4).
Les récepteurs T situés à la surface sont constitués de deux chaînes protéiques reliées par des ponts disulfures. Ils ne peuvent se lier qu'à un seul antigène.
Les LT possèdent aussi un corécepteur (CD) qui varie selon le type de LT.
Un LT ne peut pas reconnaître directement un antigène. Il reconnaît les déterminants antigéniques portés par des cellules présentatrices d'antigènes ou par des cellules parasitées par un virus. De ce fait, les LT doivent non seulement reconnaître l'antigène mais aussi la cellule qui le porte. C'est le rôle du système HLA ou CMH (complexe majeur d'histocompatibilité).
Celui-ci est mis en place par une série de gènes. Il existe deux catégories de CMH, le CMH de classe I et la CMH de classe II, selon le type de cellules.
Dans le cadre d'une première approche des mécanismes d'immunité, nous ne rentrerons pas dans ces distinctions subtiles. On considérera le CMH comme un tout, selon le schéma suivant qui présente donc la double reconnaissance (CMH et antigène) :
Les premières étapes de la réponse immunitaire sont les mêmes que pour les LB : sélection clonale, prolifération clonale et différenciation.
Les LT CD8 se différencient en lymphocytes T cytotoxiques et en LT CD8 mémoire assez peu efficaces.
Les LT cytotoxiques vont se fixer sur les cellules parasitées par l'antigène grâce à une double reconnaissance, celle du CMH et celle des déterminants antigéniques portés à la surface des cellules parasitées. Les LT cytotoxiques vont alors les détruire.
Les LT cytotoxiques peuvent provoquer soit l'autodestruction de la cellule par apoptose, soit l'éclatement de la cellule par choc osmotique.
Les LT CD4 se différencient en LT auxiliaires et en LT CD4 mémoire.
Les LT auxiliaires sécrètent de l'interleukine 2 qui permet la différenciation des LB et des LT. Ils constituent le pivot de la réponse immunitaire adaptative.
Le VIH, un virus particulier
L'épidémie de SIDA (syndrome d'immunodéficience acquise) a commencé au débuts des années 1980. Les chiffres les plus récents sont données dans le tableau suivant :
Le mode de contamination par le VIH (virus de l'immunodéficience humaine) est essentiellement sexuel. Le SIDA est une IST (infection sexuellement transmissible).
C'est en 1983 que le virus du SIDA a été décrit pour la première fois. L'électronographie ci-dessous date de 1985.
Le VIH est un rétrovirus, c'est à dire que son matériel génétique est constitué de 2 molécules d'ARN. Son enveloppe est constituée de protéines qui sont les déterminants antigéniques du virus. A côté de l'ARN, la capside protéique contient aussi une enzyme très importante, la transcriptase-inverse.
C'est grâce aux protéines gp120 et gp41 que le virus se fixe sur les récepteurs CD4 des cellules. Ce sont donc essentiellement les LT CD4 qui sont la cible du VIH.
Lorsque le virus est fixé sur le LT CD4, sa membrane fusionne avec celui de la cellule et il injecte son ARN dans celle-ci. Grâce à la transcriptase-inverse, cet ARN est rétro-transcrit en ADN et s'intègre dans le génome du lymphocyte. L'ADN viral (appelé provirus) dirige alors le métabolisme de la cellule vers la fabrication de nouveaux virus jusqu'à destruction du lymphocyte.
Les médicaments actuels bloquent le VIH au stade provirus et l'empêche de fonctionner. On est actuellement dans l'impossibilité d'éliminer le virus.
Lorsque le virus pénètre dans l'organisme, il entraîne la fabrication d'anticorps anti-VIH par les LB. Il faut un certain temps entre le moment de l'infection et la possibilité de détecter les anticorps. C'est la fenêtre sérologique.
Le test de détection des anticorps est appelée ELISA (enzyme-linked immunosorbent assay). L'ELISA le plus courant est le test en sandwich qui est présenté ci-dessous.
Lorsque le test est négatif, on considère que la séronégativité est avéré et un autre test n'est pas nécessaire. En revanche, lorsque l'ELISA donne une séropositivité, on considère qu'il y a 10% de chances que la positivité ne soit pas due à la présence d'anticorps anti-HIV. On procède alors à un test plus précis et plus complexe, le Western Blot, qui met en évidence la présence de plusieurs types d'anticorps.
Le développement de la maladie se fait en trois phases :
— la phase de primo-infection, marquée par une forte augmentation de la charge virale, puis une chute due à la réaction de l'immunité du malade (notamment les LT cytotoxiques).
— la phase asymptomatique ou de latence clinique qui peut durer plusieurs années. Seules quelques infections marquent cette période. Cependant la baisse progressive du nombre de LT CD4 et l'augmentation concurrente de la charge virale amène vers la troisième phase.
— la phase symptomatique ou de SIDA déclaré est marquée par l développement de maladies opportunistes qui profitent de l'effondrement des défenses immunitaires du patient. Celui-ci meurt d'épuisement au bout de quelques mois.
Les maladies opportunistes